Thème de recherche n°1 : Les facteurs de la famille ABC-F
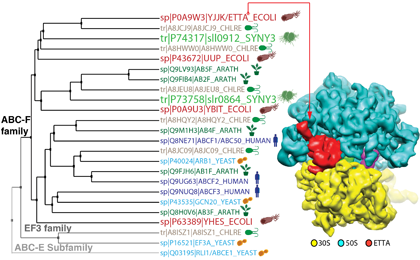
La famille ABC-F est une famille de protéines solubles, qui appartient à la superfamille des protéines ABC (ATP Binding Cassette). Les protéines qui composent cette superfamille sont des ATPase formées de domaines ABC homologues. La plupart des protéines ABC sont des transporteurs membranaires, mais la famille ABC-F est caractérisée par des protéines solubles. Cette famille est la plus commune des protéines solubles au sein de la superfamille des protéines ABC conservée des bactéries à l'homme. Quatre représentants de cette famille sont présents chez Escherichia coli, deux chez la cyanobactérie Synechocystis, six chez Chlamydomonas reinhardtii (dont un chloroplastique), cinq chez les plantes, et trois chez l'homme. Avec nos collaborateurs, nous avons démontré que le représentant prédominant de la famille ABC-F présent chez les eubactéries, la protéine EttA (energy-sensing translational throttle A) est un facteur de traduction qui régule l'entrée du ribosome dans le cycle d'élongation en fonction du ratio ADP/ATP.
En présence d'ADP, EttA inhibe la formation de la première liaison peptidique et bloque ainsi l'entrée du complexe de démarrage dans le cycle d'élongation de la traduction. La conformation d'EttA liée à l'ATP stabilise le complexe d'initiation dans une conformation favorable à la formation de la première liaison peptidique. Cette interaction stimule l'hydrolyse de l'ATP par EttA, ce qui conduit à sa dissociation du ribosome et permet au facteur EF-G de catalyser le premier évènement de translocation et ainsi l'entrée dans le cycle d'élongation. In vivo, les cellules pour lesquelles le gène ettA est inactivé, ont un défaut d'aptitude à survivre une longue phase stationnaire lorsqu'elles sont en co-culture avec les cellules sauvages. Il est connu que durant la phase stationnaire, le niveau d'ATP intracellulaire est faible.
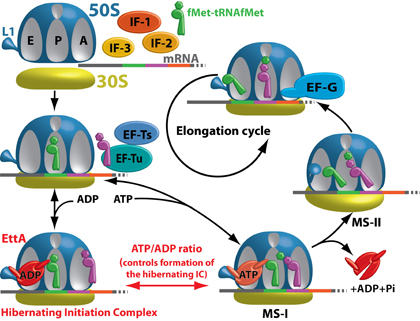
Comme la traduction de l'ARNm en protéine consomme plus de la moitié de l'énergie cellulaire, il n'est donc pas surprenant de trouver un facteur de traduction qui régule ce processus. Maintenant, nous étendons notre étude aux 3 autres paralogues d'EttA présents chez E. coli et à des orthologues d'EttA responsables de la résistance aux antibiotiques chez plusieurs bactéries pathogènes. Nous étudions aussi la fonction des deux protéines ABC-F présentes chez la cyanobactérie Synechocystis. Notre objectif est de cartographier les différentes actions des protéines ABC-F sur l'appareil de traduction.
Thème de recherche n°2 : Identification et caractérisation fonctionnelle de nouveaux facteurs de traduction putatifs
Pour ce deuxième projet, nous allons développer une méthodologie de criblage pour identifier le phénotype lié à des défauts de traduction dans une collection de mutants. Cette collection sera composée de mutants de délétion de protéine connue pour interagir avec le ribosome et dont la fonction reste non définie. En parallèle, nous allons tenter une approche par « pull-down » pour identifier de nouveaux facteurs qui interagissent avec les ribosomes bloqués durant la traduction de l'ARNm.